 题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[主观题]
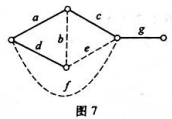
图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T。(1)求G对应T的所有基本回路。(2)求G
图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T。(1)求G对应T的所有基本回路。(2)求G
图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T。
(1)求G对应T的所有基本回路。
(2)求G对应T的所有基本割集。
查看答案

 如果结果不匹配,请 联系老师 获取答案
题目内容
(请给出正确答案)
如果结果不匹配,请 联系老师 获取答案
题目内容
(请给出正确答案)
图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T。
(1)求G对应T的所有基本回路。
(2)求G对应T的所有基本割集。
如果结果不匹配,请 联系老师 获取答案
 更多“图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T…”相关的问题
更多“图7中所示的无向图G中,实线边所表示的子图为G的一棵生成树T…”相关的问题
无向图G如图16.26所示,其中实线边为G的一棵生成树T。
(1)求G对应T的基本回路系统。
(2)求G对应T的基本割集系统。
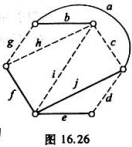
存储器的地址空间分布图和存储器的地址译码电路分别如图4.17(a)和(b)所示,图(b)中的A、B两组跨接端子可按要求分别进行接线,如1~4中的任一端子可以和5~7中的任一端子跨接。74139是2线—4线译码器(译码输出低电平有效),使能端G接地表示译码器处于正常译码状态。试完成A组跨接端子与B组跨接端子内部的连接,以便使地址译码电路按图(a)的要求进行正确寻址。
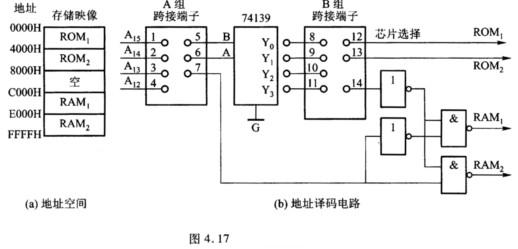
求如图7-30所示连通图G的生成树TG.设有如下“破圈法”:

(1)令G=G1,i=1;
(2)若Gi无环,则TG=Gi,否则进入(3);
(3)在Gi中找出一个环σi,并从中删去边ei,令Gi+1=Gi-ei;
(4)i=i+1,返回(2).
设有如图8—2所示的程序流图G:
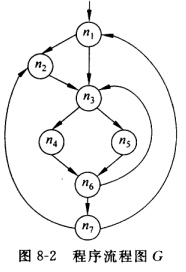 (1)给出G中如下结点的必经结点集: D(6)={ } D(7)={ } (2)给出G中所有回边和循环。
(1)给出G中如下结点的必经结点集: D(6)={ } D(7)={ } (2)给出G中所有回边和循环。
A.G'为G的子图
B.G'为G的连通分量
C.G'为G的极小连通子图且V'=V
D.G'是G的无环子图
对于一个具有n个顶点和e条边的无向图,若采用邻接表表示,则表头向量的大小为( ),所有邻接表中的结点总数为( )。
由MNNG(亚硝基胍)引起的诱变损伤的本质以及它从DNA上被修复的机制可以用下面的实验来鉴定。为了确定诱变损伤的本质,未经处理的细菌和已用低剂量MNNG处理的细菌都在含50μg/ml的3H-MNNG的培养物中培养10min。分离它们的DNA并水解成核苷酸,然后经过纸层析分析放射性的嘌呤,结果如图Q12.2所示:

图Q12.2 层析法分离未被处理和已被低剂量MNNG处理的细菌DNA中被标记的甲基化嘌呤实线表示未被处理细菌DNA中的甲基化嘌呤;虚线表示MNNG处理的细菌所得结果
为了研究诱变损伤切除的机制,首先纯化负责切除的酶,把不同量的酶(相对分子质量19000)和已被3H标记含0.26pmol突变碱基的DNA一起温育,分析切除动力学。在不同时间取样,分析DNA以确定还存在多少突变残基(图Q12.3)。当在5℃而不是37℃时重复这个实验时,虽然最初的切除速率较慢,却得到一样的终点。

图Q12.3纯化的甲基转移酶把3H标记的甲基从DNA上切除所示为纯化酶的量
某计划进度与实际进度横道图如图6_3所示,图中粗实线表示计划进度(上方的数据表示每周计划投资),粗虚线表示实际进度(上方的数据表示每周实际投资),假定各分项工程每周计划完成的工程量相等。
| 分项 工程 | 进度计划 | |||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| A | 5 | 5 | 5 | |||||||||
| (5) | (5) | (5) | ||||||||||
| 5 | 5 | 5 | ||||||||||
| B | 4 | 4 | 4 | 4 | 4 | (4) | ||||||
| (4) | (4) | (4) | (4) | |||||||||
| 4 | 4 | 4 | 4 | 4 | ||||||||
| C | 9 | 9 | 9 | 9 | (9) | (9) | ||||||
| (9) | (9) | |||||||||||
| 8 | 7 | 7 | 7 | |||||||||
| D | 5 | 5 | 5 | 5 | (4) | (4) | ||||||
| (4) | (4) | (4) | ||||||||||
| 4 | 4 | 4 | 5 | 5 | ||||||||
| E | 3 | 3 | 3 | (3) | (3) | |||||||
| (3) | ||||||||||||
| 3 | 3 | 3 | ||||||||||
某计划进度与实际进度横道图

















