 题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[主观题]
图12-3-3示两个同源DNA结构。在每个分子中(+)链互补于其本身的(-)链以及另一分子的(-)链。分子的排列方向是两
图12-3-3示两个同源DNA结构。在每个分子中()链互补于其本身的(-)链以及另一分子的(-)链。分子的排列方向是两个b端的碱基顺序同源。上面的分子比下面的分子短,少了一段a端(见图12-3-3)。使分子杂交并发生分支移动。结果会得到图中(1)至(6)中的哪种结构?

查看答案

 如果结果不匹配,请 联系老师 获取答案
如果结果不匹配,请 联系老师 获取答案

 更多“图12-3-3示两个同源DNA结构。在每个分子中(+)链互补…”相关的问题
更多“图12-3-3示两个同源DNA结构。在每个分子中(+)链互补…”相关的问题



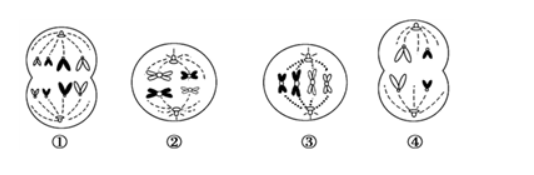



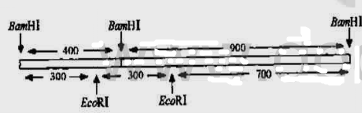 图025.1杂合基因的结构
图025.1杂合基因的结构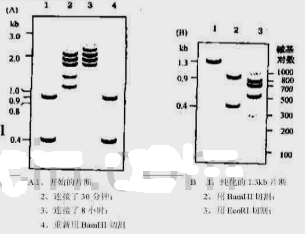 图Q25.2纯化DNA片段连接后电泳检测
图Q25.2纯化DNA片段连接后电泳检测















