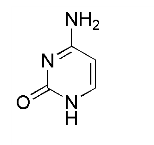
题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[主观题]
假定你制取了两个DNA样品,一个正常,另一个变性,各用0.01mol/L NaCl溶液透析过。透析前并不知道A260值。然后,
假定你制取了两个DNA样品,一个正常,另一个变性,各用0.01mol/L NaCl溶液透析过。透析前并不知道A260值。然后,每个样品中各加入少量酶,这时,两个样品搞混了。如何用吸收值来区别这两个样品?假定检测中用去了一小部分样品,且酶不会干扰测定。再设计第二种检测方法,确定它不需引入其他试剂或引起变性的条件。
查看答案

 如果结果不匹配,请 联系老师 获取答案
如果结果不匹配,请 联系老师 获取答案

 更多“假定你制取了两个DNA样品,一个正常,另一个变性,各用0.0…”相关的问题
更多“假定你制取了两个DNA样品,一个正常,另一个变性,各用0.0…”相关的问题