 题目内容
(请给出正确答案)
题目内容
(请给出正确答案)
[多选题]
Takara 专门的DNA纯化产品可以分为()
A.DNA片段纯化
B.质粒DNA纯化
C.基因组DNA纯化
D.线粒体DNA
 答案
答案
D质粒DNA纯化基因组DNA纯化
 如果结果不匹配,请 联系老师 获取答案
题目内容
(请给出正确答案)
如果结果不匹配,请 联系老师 获取答案
题目内容
(请给出正确答案)
A.DNA片段纯化
B.质粒DNA纯化
C.基因组DNA纯化
D.线粒体DNA
答案
D质粒DNA纯化基因组DNA纯化
如果结果不匹配,请 联系老师 获取答案
 更多“Takara 专门的DNA纯化产品可以分为()”相关的问题
更多“Takara 专门的DNA纯化产品可以分为()”相关的问题
片段重新连接起来,得到如图Q25.1所示的结果。
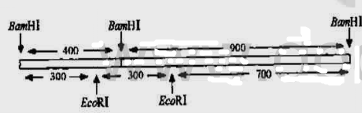 图025.1杂合基因的结构
图025.1杂合基因的结构
于是将这两种片段混合起来,并在连接酶的存在下进行温育,分别在30min和8h取样进行凝胶电泳分析。令人惊讶的是,连接产物并非是理想中的1.3kb的重组分子,而是一种复杂的片段模式[图Q25.2(a)]。同时发现随着温育时间的延长,较小片段的浓度逐渐降低,大片段的浓度逐渐增加。如果用BamH Ⅰ来切割连接后的混合物,则起始的片段可以重新产生[图Q25.2(a)]。
从凝胶中分离纯化出1.3kb的片段,并取出一部分用BamH Ⅰ进行切割,以检查它的结构。正如预料的一样,出现了两个原始的带[图Q25.2(b)]。但是用EcoR Ⅰ切割另一部分样品,希望能够产生两个300bp和一个长度为700bp的核苷酸片段,然而,凝胶电泳的结果,令人惊奇[图Q25.2(b)]。
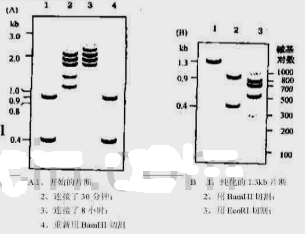 图Q25.2纯化DNA片段连接后电泳检测
图Q25.2纯化DNA片段连接后电泳检测
因子通常已经失活。你可以想出一个极好的方法以加快分析速度。你可以合成一段不含胞嘧啶核苷酸的DNA序列,将这段序列放到启动子后在一适当反应混合物中温育,这个启动子将指导不含鸟嘌呤核苷酸的转录本的合成,如果不加GTP,只产生一定长度的RNA转录本,这个RNA转录本来自于合成的DNA序列。因为其他转录本都在需G处被终止。如果你能使这个想法得到实现,则快速分析特殊序列的转录仅通过测量标记核苷酸的放射性就可实现。
为了验证你的想法,你构建两个携带合成DNA的质粒。一个具有腺病毒的启动子(PmL1),另一个无启动子(PC1)。把两种质粒同纯化的RNApolⅡ、纯化好的转录因子样品及32P-CTP混合,除此之外,再加入各种组合的GTP、RNaseT1(可以切开RNA与G的连接)和3'-氧甲基GTP(无论何时它参入正在形成的链,它就会终止转录),通过凝胶电泳测定结果如图13-3-42所示。
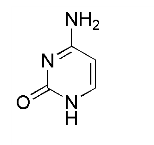
在真核细胞中DNA折叠成为染色质是如何影响转录的?你可以应用C-负转录单元解决这个问题。这个模板在RNApolⅡ及四个转录因子(TFⅡA、TFⅡB、TFⅡD和TFⅡE)存在下能很好地转录。
为了检验染色质对转录的影响,首先将模板装配成核小体(核小体来自蛙卯母细胞抽提物)并纯化染色质模板,然后加入转录组分,结果没有发生转录(图13-3-48)。然后改变步骤,首先,在缺少NTPs的情况下将模板和转录组分一起温育,然后将模板装配为核小体,再纯化染色质模板。现在,当要再加入转录组分时,转录顺利进行就如在裸露的DNA模板上进行的转录一样,这样得出结论:一个或多个转录组分一定同模板结合,然后使启动子可以接近。

为了更进一步了解这种现象,再做两个附加实验。在第~个实验中,在温育过程中少加一种转录组分;在第二个实验中,在进行转录分析中少加一种转录组分。
你纯化了与这种效应有关的蛋白,显示它使非缺失的模板及-61缺失的模板转录都增强了10倍,面对-50缺失的模板无激活作用,印迹分析表明这个因子同一个TATA位点上游的特殊的短序列相结合,虽然这种因子在TFⅡD缺乏时同它的结合位点的结合是短暂的,但当TFⅡD存在时,它可与结合位点稳定地结合。

利用一个包含纯化的RNA聚合酶的核心酶、DNA模板、σ因子以及标记核苷酸(32pppN)的系统,来研究σ因子在原核转录中的作用。在10-10mol和10-11mol两种不同浓度的核心酶催化下32P掺入到RNA的图如下所示,σ因子浓度恒定为10-12mol。

A.帐户名和口令认证方式是计算机身份认证中最常用的认证方式
B.消息认证必须有专门的硬件支持才可以实现
C.认证技术可以识别所访问的 IP地址是否合法
D.DNA认证是目前计算机身份认证方式中最常用的认证方式
你纯化出经BamHI消化重组质粒DNA所得的两个DNA片段,其中一个长为400bp,另一个长900bp,你想要如图16-3-20(1)所示将它们连接起来产生一个杂合基因,如果你的推测是正确的话,就会得到令人惊奇的新物质。

你在DNA连接酶存在情况下将两种片段混合在一起并且进行孵育,30分钟后和8小时后,你取一些样品进行琼脂糖凝胶电泳分析,你惊讶地发现连接得到了一系列复杂的DNA片段而不是所需的1.3kb小片段,并且发现孵育时间越长,小片段浓度越低而大片段浓度增加,如果你用BamHI切割连接后的重组体,你会重新获得开始的两片段(图16-3-20(2)A)。

令人吃惊的是,你将从琼脂糖凝胶中纯化得到的1.3kb片段用BamHI消化,检查它的结构,与预想的一样,可以得到最初的两片段(图16-3-20(2)B)。这就可以确定这是你所需的结构。但是,你用EcoRl消化另一份样品,你希望通过消化得到300bp的片段和700bp的片段,但你跑的电泳是一系列DNA片段(图16-3-20(2)B)。
由MNNG(亚硝基胍)引起的诱变损伤的本质以及它从DNA上被修复的机制可以用下面的实验来鉴定。为了确定诱变损伤的本质,未经处理的细菌和已用低剂量MNNG处理的细菌都在含50μg/ml的3H-MNNG的培养物中培养10min。分离它们的DNA并水解成核苷酸,然后经过纸层析分析放射性的嘌呤,结果如图Q12.2所示:

图Q12.2 层析法分离未被处理和已被低剂量MNNG处理的细菌DNA中被标记的甲基化嘌呤实线表示未被处理细菌DNA中的甲基化嘌呤;虚线表示MNNG处理的细菌所得结果
为了研究诱变损伤切除的机制,首先纯化负责切除的酶,把不同量的酶(相对分子质量19000)和已被3H标记含0.26pmol突变碱基的DNA一起温育,分析切除动力学。在不同时间取样,分析DNA以确定还存在多少突变残基(图Q12.3)。当在5℃而不是37℃时重复这个实验时,虽然最初的切除速率较慢,却得到一样的终点。

图Q12.3纯化的甲基转移酶把3H标记的甲基从DNA上切除所示为纯化酶的量
A.必须分离纯化出个体染色体。
B.DNA必须仅从单倍体细胞(基因组)分离,这可避免由于二倍体细胞中同源染色体的存在而引起的模糊信息。
C.必须进行单个染色体分析,这样的分析仅在细胞分裂后期才有可能进行。
D.必须发现一些在染色体上仅有一个切割位点的限制酶。
E.必须先分离核DNA和细胞器DNA,然后用它们来分析个体的图谱。

















